【泛基因家族一区】8个油菜 SPL 泛基因家族分析(泛基因组构建策略)
一、文章基本信息
- 在线发表时间: 2025年9月15日
- 第一单位:湖南农业大学农学院
- 通讯作者: Zhongsong Liu
- 文章题目: 甘蓝型油菜SPL基因家族的泛基因组分析及其在种子重量和产量中的功能探索
- 发表期刊: 《工业作物与产品》(Industrial Crops & Products)

二、摘要
提高油菜产量是育种者的关键目标,而种子重量是提升油菜品种产量的关键因素。针对种子重量的研究日益关注SQUAMOSA启动子结合蛋白样(SPL)家族,使得SPL基因成为我们研究的主要目标。我们对八个油菜品种进行了全基因组分析,鉴定出72个BnaSPL基因,其中包括53个核心基因、14个非必需基因和5个特有基因。基于进化关系,这些BnaSPL基因被分为六个亚组。对Ka/Ks值的评估显示,7个BnaSPL基因受到正选择,而44个受到纯化选择。不同组织的表达谱分析和qRT-PCR分析证实,26个BnaSPL基因在种子发育过程中高表达。进一步分析表明,八个基因在至少两个油菜品种中存在结构和基序的变异,尽管它们都保留了保守的SBP结构域。顺式作用元件的多样性提示了这些基因潜在的功能变异。对2311份种质资源的单倍型分析确定了四个关键基因——BnaSPL2–3、BnaSPL3–3、BnaSPL7–4和BnaSPL16–4,其单倍型与不同油菜品种间的千粒重和产量差异相关。值得注意的是,这些产量提升是稳定的,并且不受外部环境因素(如低或高盐胁迫)的影响。我们的发现强调了BnaSPL2–3、BnaSPL3–3、BnaSPL7–4和BnaSPL16–4在培育具有更高千粒重和产量的油菜品种中的潜力。

三、结果
图1:72个BnaSPL基因与17个AtSPL基因的系统发育树
主要内容: 描述了基于拟南芥和甘蓝型油菜SPL基因家族成员蛋白序列构建的系统发育树。共鉴定出72个BnaSPL基因和17个AtSPL基因,并将其分为六个组(I-VI)。每组都至少包含一个拟南芥和一个油菜SPL蛋白,表明SPL蛋白的主要特征在拟南芥和油菜分化前就已确立。其中,第VI组是最大的,包含36个SPL基因。值得注意的是,拟南芥的AtSPL12和AtSPL13A在甘蓝型油菜中没有直系同源基因,表明这些基因在油菜进化过程中丢失了。系统发育树中,圆形代表核心基因,方形代表非必需基因,星形代表特有基因。
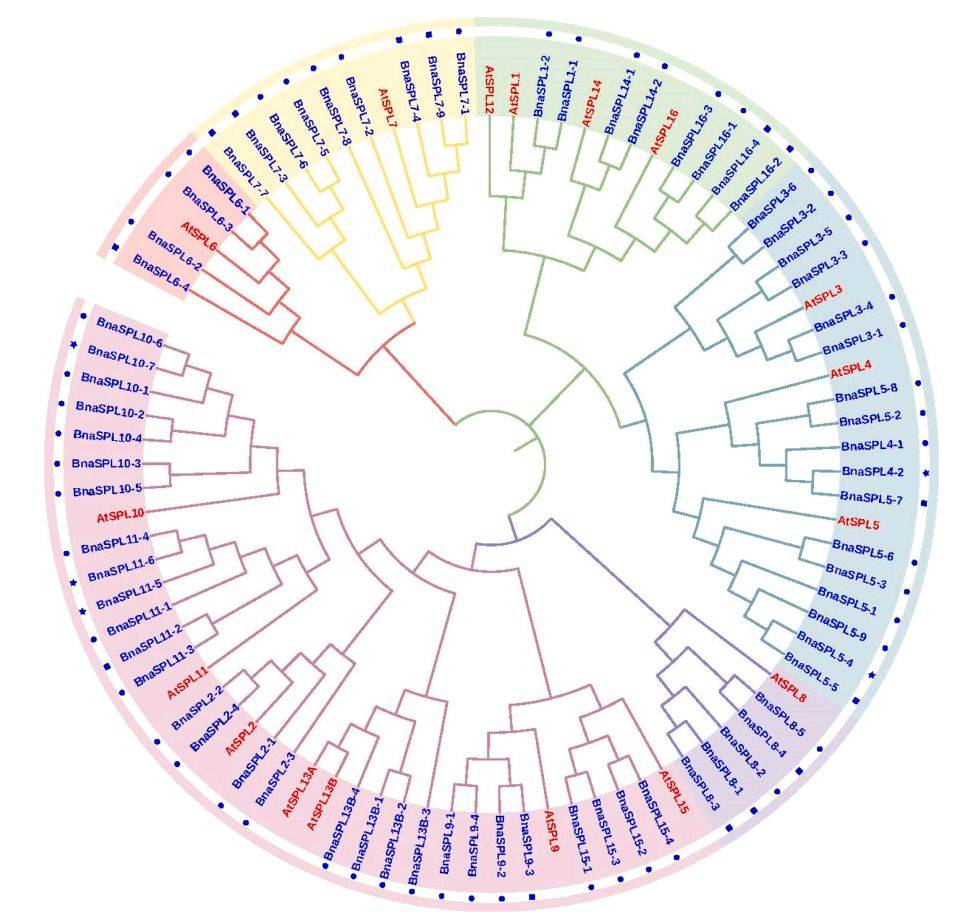
图例说明:
- 圆形代表核心基因(存在于所有分析品种中),方形代表非必需基因(存在于部分品种中),星形代表特有基因(仅存在于某个特定品种中)。

图2:八个甘蓝型油菜品种中BnaSPL基因的共线性分析
主要内容:
展示了对八个油菜品种中BnaSPL基因进行共线性分析的结果。分析显示,参考基因组ZS11与其他品种之间存在大量的同源基因对,例如与Gangan有62对,与Shengli有61对,与Zheyou有62对等。这些结果表明ZS11与Gangan、Shengli、Westar和Zheyou之间存在较高的同源性和保守关系。图2通过多重共线性图可视化了这些同源关系,揭示了不同油菜基因组间BnaSPL基因的进化保守性和潜在的基因组重排事件。

图例说明:
- 每条彩色曲线连接一对同源的BnaSPL基因,表明它们起源于共同的祖先基因。不同颜色可能代表不同的染色体或基因组区块。结果显示参考基因组ZS11与其他几个品种(如Gangan、Shengli)之间存在大量保守的同源关系。


图 3 : 八个甘蓝型油菜品种中BnaSPL基因的Ka/Ks值分布
主要内容:
图3分析了BnaSPL基因在不同油菜品种间的Ka/Ks值分布,以探究其进化选择压力。Ka/Ks值用于衡量非同义替换与同义替换的比率,以推断基因所受的选择类型。分析发现,大多数BnaSPL基因的Ka/Ks峰值在0到1之间,表明这些基因主要经历了纯化选择,以去除有害突变。然而,有7个基因(如BnaSPL1–3、BnaSPL5–8等)在特定品种中显示出Ka/Ks值大于1,表明它们可能受到了正选择,在进化过程中适应性发生了变化。图3以山脊线图的形式展示了所有BnaSPL基因在八个品种中的Ka/Ks值分布情况,直观地反映了不同基因所承受的选择压力差异。
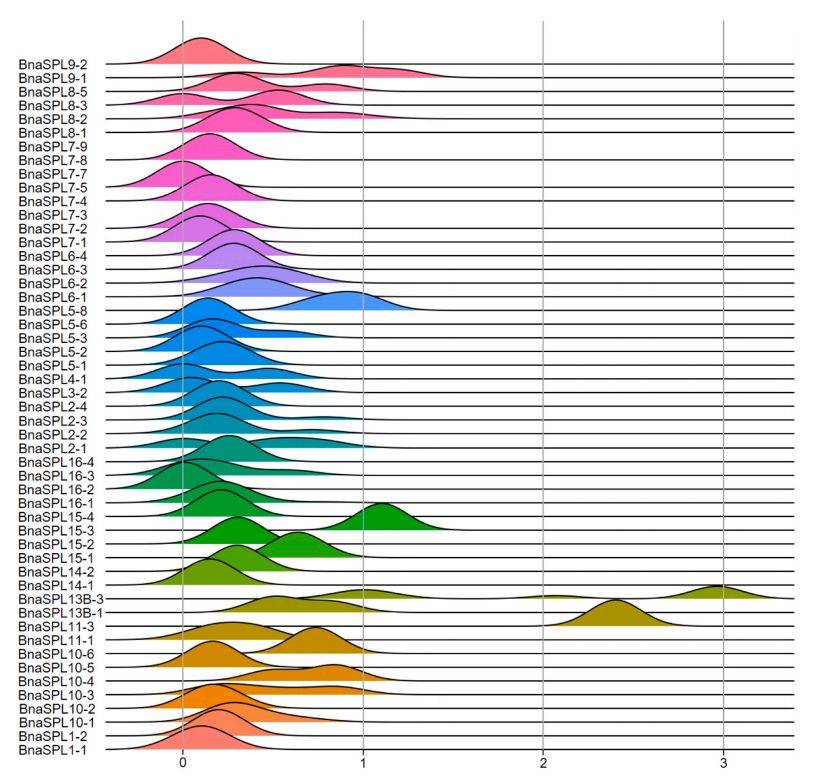
图例说明:
- 每条“山脊”代表一个BnaSPL基因,其峰值的水平位置(X轴)表示Ka/Ks值的大小。峰值位于0-1区间(左侧)表示基因主要受纯化选择;峰值大于1(右侧)则表示在特定比较中受到了正选择。图形直观显示大部分基因受纯化选择,少数基因显示了正选择的信号。

图 4 :甘蓝型油菜中BnaSPL基因的表达谱
主要内容:
图4研究了BnaSPL基因的表达模式。通过分析根、茎、叶、芽、花、角果和种子等不同组织的转录组数据(图4A热图),发现一些BnaSPL基因在所有组织中均高表达(如BnaSPL1、BnaSPL6等),而另一些则呈现组织特异性表达(如BnaSPL10–4在茎中高表达)。特别关注种子发育,分析了29个在种子中高表达的BnaSPL基因在不同发育阶段(14-64天)的表达变化(图4B热图)。进一步通过qRT-PCR在开花后20天和40天的种子中验证了这29个基因的表达(图4C柱状图),结果与RNA-seq数据基本一致,最终确认有26个BnaSPL基因在种子发育期持续高表达,提示它们可能在调控种子重量中发挥作用。
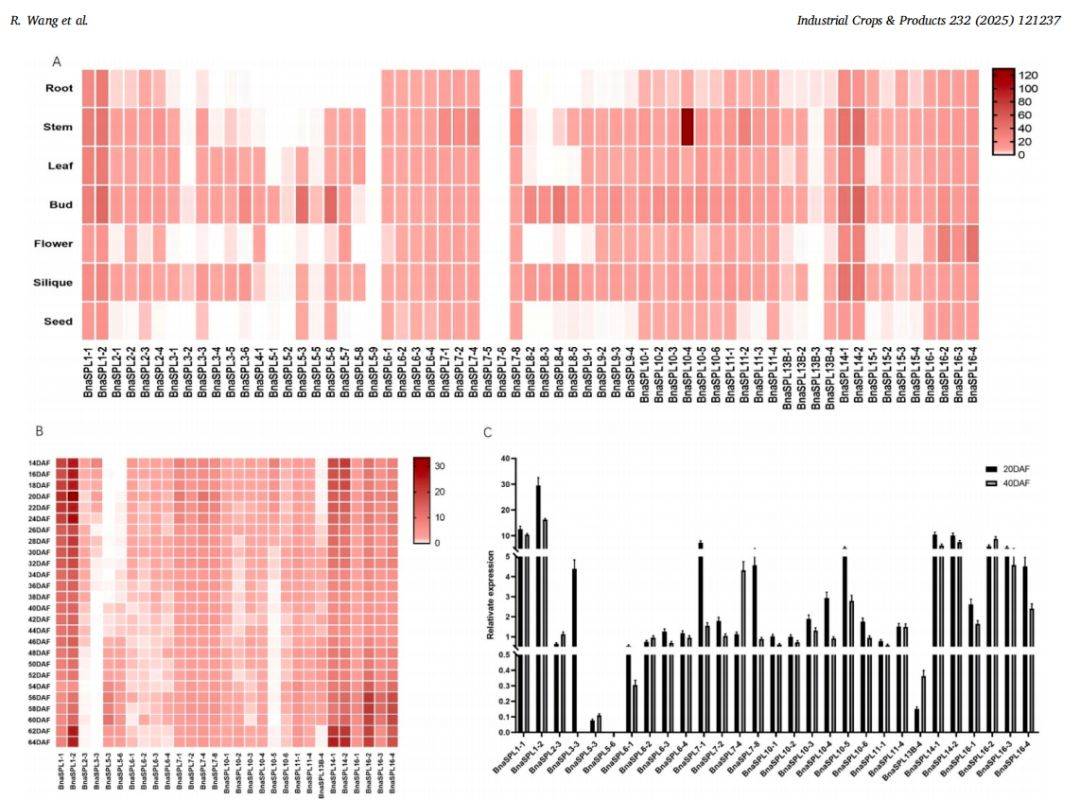
图例说明:
- A. 不同组织中BnaSPL基因的表达热图: 热图显示了BnaSPL基因在根、茎、叶、芽、花、角果和种子七种组织中的表达水平。颜色从蓝到红表示表达量从低到高。结果显示部分基因组成型高表达,部分基因呈现组织特异性表达。
- B. 种子发育过程中29个BnaSPL基因的表达热图: 热图聚焦于在种子中高表达的29个基因,展示了它们在开花后14至64天不同发育阶段的表达动态。颜色深浅代表相对表达量。
- C. 29个BnaSPL基因在开花后20天和40天种子中的qRT-PCR表达验证: 柱状图通过实验方法(qRT-PCR)验证了热图(B)中29个基因在种子两个关键发育时期的表达量。柱高表示相对表达量,误差线代表标准差。结果与转录组数据基本吻合,确认了26个基因在种子发育期持续高表达。

图 5 :八个油菜品种中BnaSPL基因的系统发育树、基因结构、基序、结构域及顺式作用元件比较
主要内容:
图5深入分析了可能调控种子重量的26个BnaSPL基因在八个油菜品种中的进化、结构、基序、结构域和顺式作用元件。图5(以及补充图S1-3)展示了代表性基因(如BnaSPL2-3、BnaSPL3-3等)的综合分析结果。研究发现,其中8个基因在不同品种间存在内含子/外显子数量或基序组成的变异。尽管存在结构变异,所有基因都保留了保守的SBP DNA🧬结合结构域,其中BnaSPL16–1和BnaSPL16–3还含有LamG和ANKRY超家族结构域。对启动子区域顺式作用元件的分析发现了与胁迫响应、激素响应以及植物生『长发』育相关的多种元件,其中光响应元件存在于所有品种中,而其他元件(如干旱诱导元件、生长素响应元件等)则表现出基因和品种特异性。这些变异,尤其在No2127和Zheyou品种中更为集中,可能与其特殊的育种方式(如远缘杂交、突变育种)有关。

图例说明:
- 左侧系统发育树: 显示该基因在进化中的位置。
- 基因结构: 黄色框代表外显子,黑色线代表内含子。不同品种间外显子数量或长度可能存在差异。
- 基序: 不同颜色的框代表不同的保守蛋白基序(Motif),数字编号不同。
- 结构域: 绿色框标注保守的SBP结构域(可能还有其他结构域如LamG)。
- 顺式作用元件: 启动子区域上不同形状的标记代表不同类型的调控元件(如光响应、激素响应、胁迫响应等)。不同品种间元件的种类和数量存在变异。
6 :BnaSPL2–3单倍型对千粒重和产量性状的影响
主要内容:
该图综合展示了BnaSPL2–3基因单倍型与性状的关联。首先通过序列变异分析确定了Hap1和Hap2两种主要单倍型。关联分析显示,携带Hap1的种质具有显著更高的千粒重。在种子发育过程中,Hap1的表达量也显著高于Hap2,表明该基因可能通过正向调控影响种子重量。更重要的是,在正常、低盐碱和高盐碱三种环境下,Hap1单倍型均能带来显著更高的单株产量,且这种增产效应不受逆境胁迫影响,证明了其优异的稳产性。
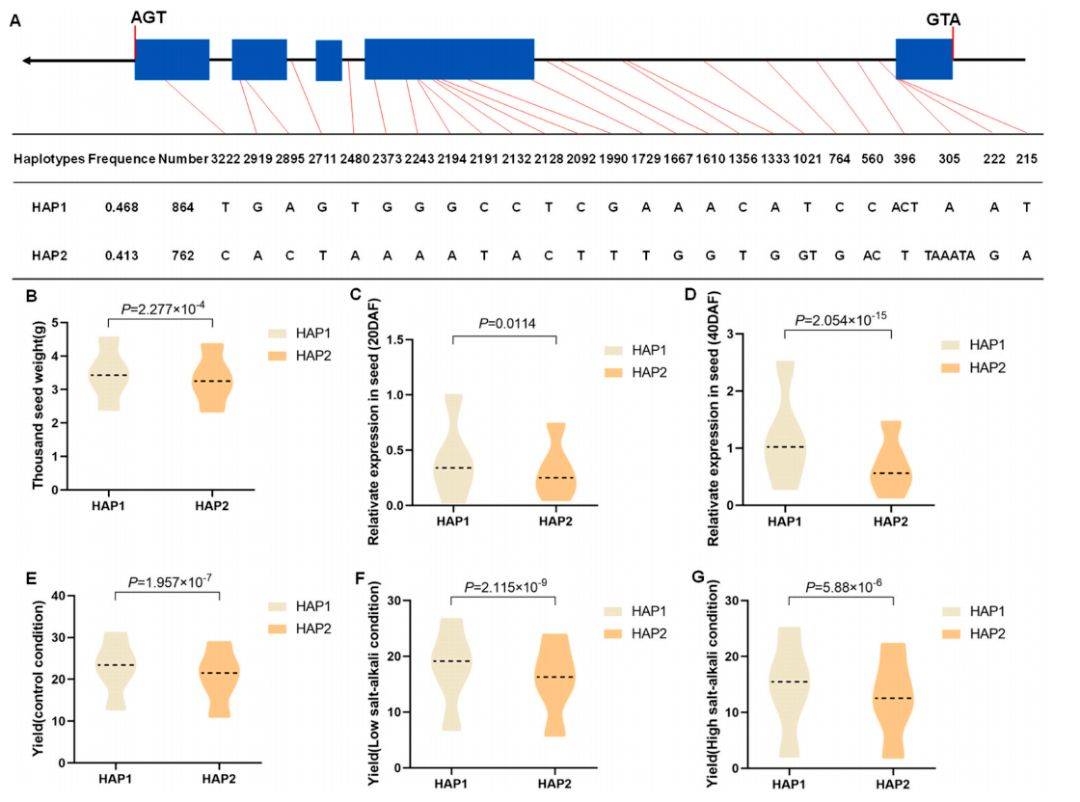
图例说明:
- A. 主要单倍型: 展示Hap1和Hap2单倍型的DNA🧬序列变异(SNP/InDel)模式。
- B. 不同单倍型材料的千粒重比较: 箱线图显示携带Hap1的材料千粒重显著高于Hap2。
- C-D. 不同单倍型在种子发育中的相对表达量: 柱状图显示在开花后20天和40天,Hap1的表达量均高于Hap2。
- E-G. 不同单倍型在对照、低盐碱和高盐碱条件下的产量比较: 箱线图显示在三种环境下,携带Hap1的材料单株产量均显著高于Hap2。
7 :BnaSPL3–3单倍型对千粒重和产量性状的影响
主要内容:
该图揭示了BnaSPL3–3基因独特的调控模式。基于序列变异定义了Hap1和Hap2单倍型,其中Hap1同样与更高的千粒重相关。然而,与BnaSPL2–3不同,在种子发育期,Hap1单倍型的表达量反而显著低于Hap2,暗示该基因可能通过一种负向调控机制(如下调表达有利于种子增重)来影响性状。尽管如此,在三种环境条件下,携带Hap1单倍型的材料其单株产量均稳定且显著高于Hap2,展现了其与BnaSPL2–3相似的、可靠的增产潜力。
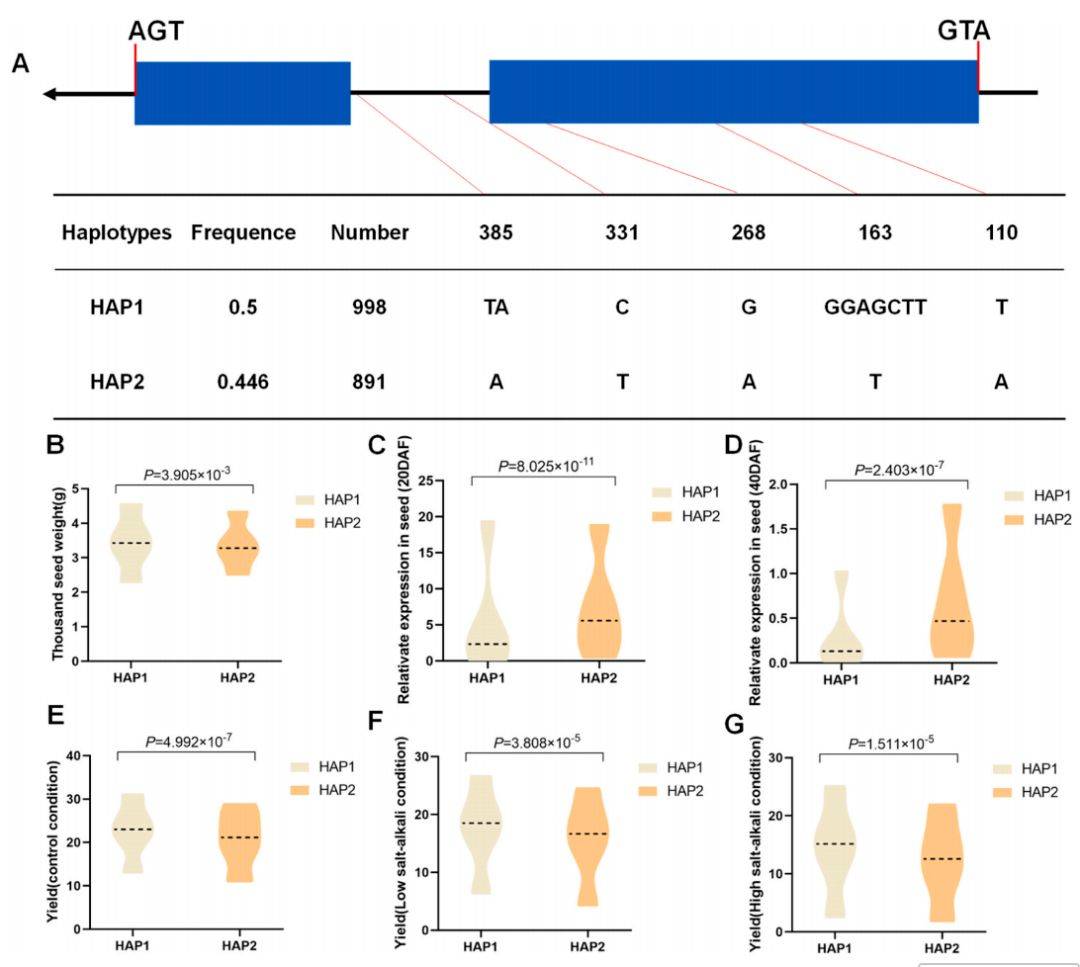
图例说明:
- B图中Hap1的千粒重更高,但C-D图中Hap1的表达量却低于Hap2,暗示其可能起负调控作用。E-G图中Hap1的产量同样更高。
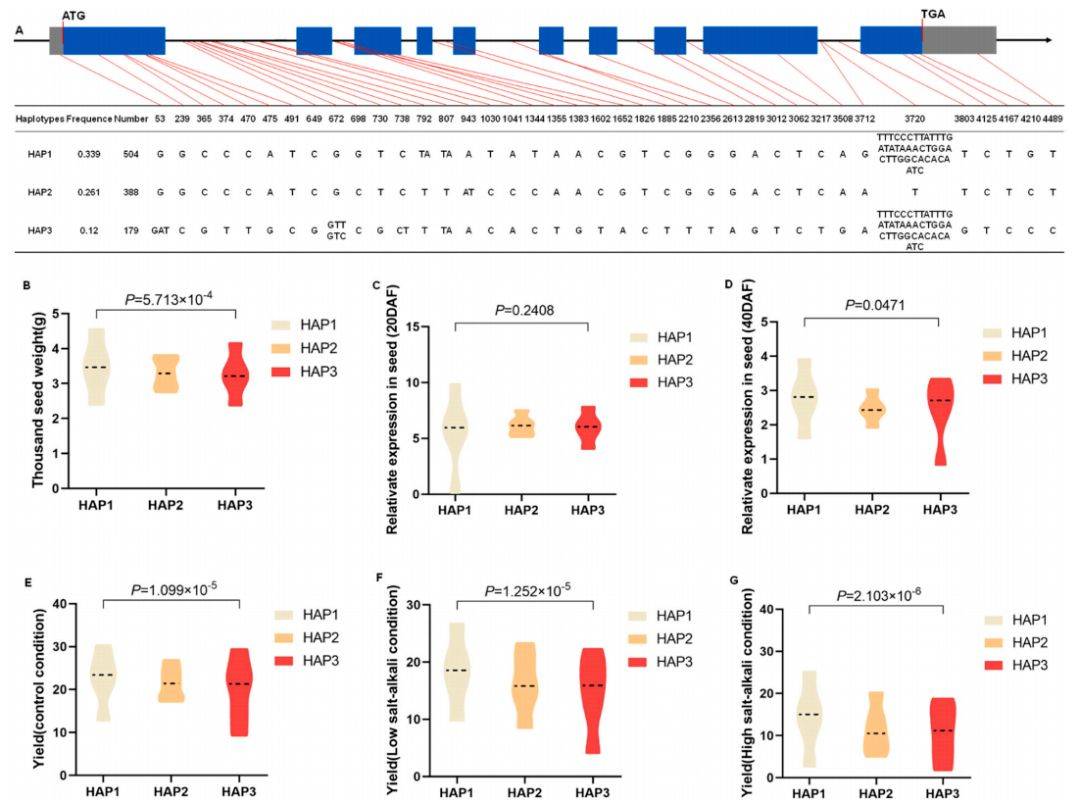
8 :BnaSPL7–4单倍型对千粒重和产量性状的影响
主要内容:
该图展示了BnaSPL7–4基因三个主要单倍型(Hap1、Hap2、Hap3)与性状的关系。其中,Hap1单倍型与最高的千粒重显著相关。值得注意的是,不同单倍型之间在种子发育期的基因表达水平并无显著差异,表明该基因的变异可能主要通过影响蛋白质功能或翻译后修饰等途径来调控性状。在产量方面,Hap1单倍型在正常与盐碱胁迫环境下均能带来显著更高的单株产量,证明了其对增产和抗逆的贡献。
图例说明:
- A图中展示了三个主要单倍型(Hap1, Hap2, Hap3)。B图显示Hap1的千粒重显著高于Hap3。C-D图中不同单倍型间表达量无显著差异。E-G图中Hap1的产量在三种环境下均具优势。
9 :BnaSPL16–4单倍型对千粒重和产量性状的影响
主要内容:
该图呈现了BnaSPL16–4基因的单倍型效应,其模式与BnaSPL7–4高度相似。在鉴定的Hap1、Hap2、Hap3三种单倍型中,Hap1与显著更高的千粒重相关联。同样,不同单倍型间的表达量没有显著差异。在产量性状上,携带Hap1单倍型的材料在多种环境条件下均表现出稳定且显著更高的单株产量,进一步确认了该单倍型在培育高产、稳产品种中的应用价值。
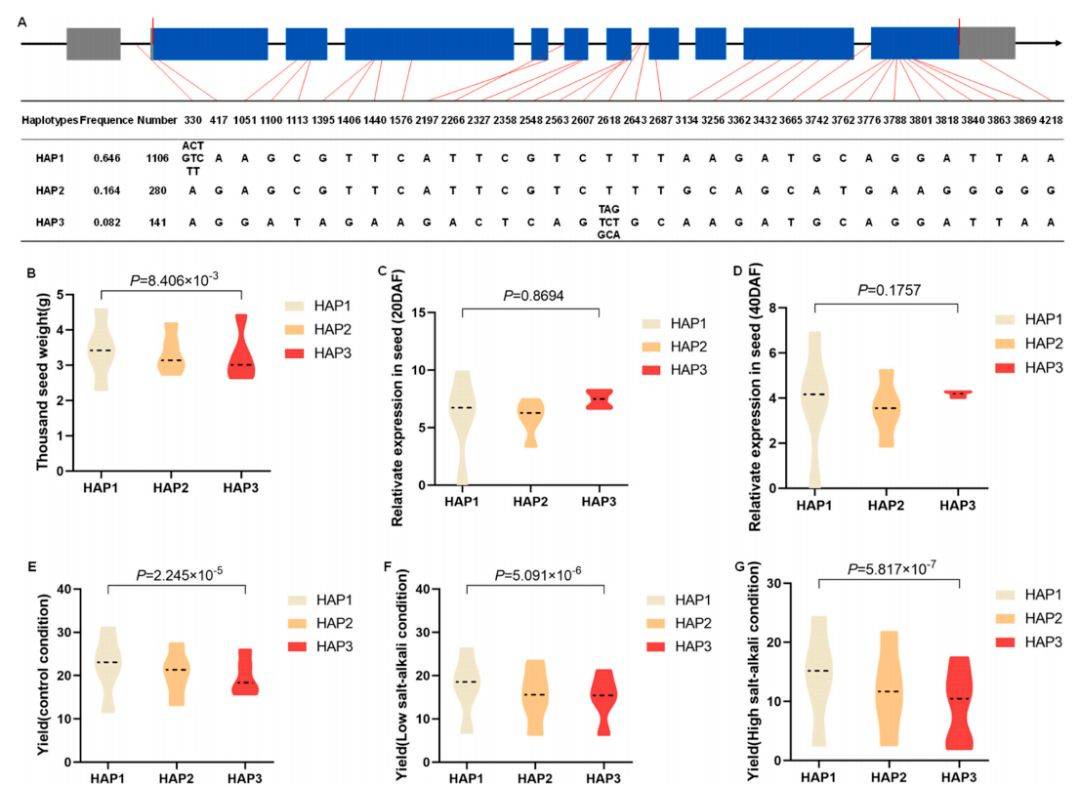
- 展示的是BnaSPL16–4基因的三个单倍型及其与性状的关联,模式与BnaSPL7–4相似。
10 :利用OrchidMD进行基因家族鉴定与CRISPR基因编辑的案例研究结果
主要内容:
讨论了另外两个基因BnaSPL10–3和BnaSPL11–1的单倍型与性状的关联。分析发现,虽然这两个基因也存在多个序列变异和单倍型(图10A, C),并且携带Hap3单倍型的材料千粒重显著高于Hap1(图10B, D),但这些单倍型与整体产量没有显著相关性。这表明,BnaSPL10–3和BnaSPL11–1可能特异性地影响种子重量,但由于单株角果数等其他产量构成因素的补偿或抵消作用,最终并未导致产量的显著变化。
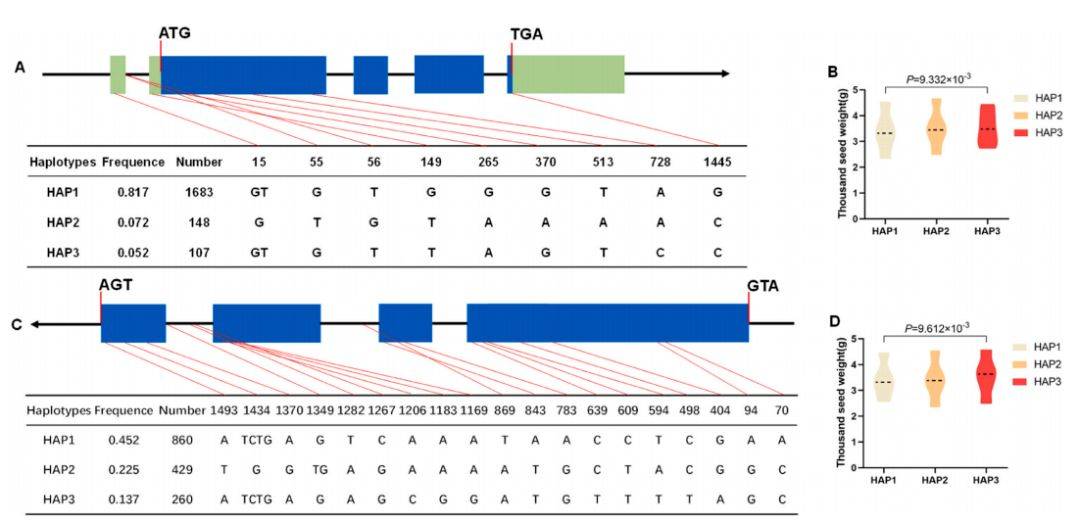
- A, C. 主要单倍型: 分别展示BnaSPL10–3和BnaSPL11–1基因的三个主要单倍型(Hap1, Hap2, Hap3)的变异模式。
- B, D. 不同单倍型材料的千粒重比较: 箱线图分别显示,对于这两个基因,携带Hap3的材料的千粒重显著高于Hap1。
11 :利用OrchidMD进行基因家族鉴定与CRISPR基因编辑的案例研究结果
主要内容:
利用千粒重极端材料(大粒品系D3和小粒品系X10)对生物信息学分析结果进行了实验验证。表型观察证实D3的种子明显大于X10(图11A),千粒重也显著更高(图11B)。对BnaSPL2–3和BnaSPL3–3的基因序列分析显示(图11C-D),D3均携带与高产相关的Hap1单倍型,而X10携带Hap2单倍型,这与前期基于大量种质的生物信息学关联分析结果完全一致,进一步证实了这两个基因在调控油菜千粒重中的关键作用。而对于BnaSPL7–4和BnaSPL16–4,在极端材料中的验证结果与群体分析不完全一致,提示其作用机制可能更为复杂或受其他因素影响。
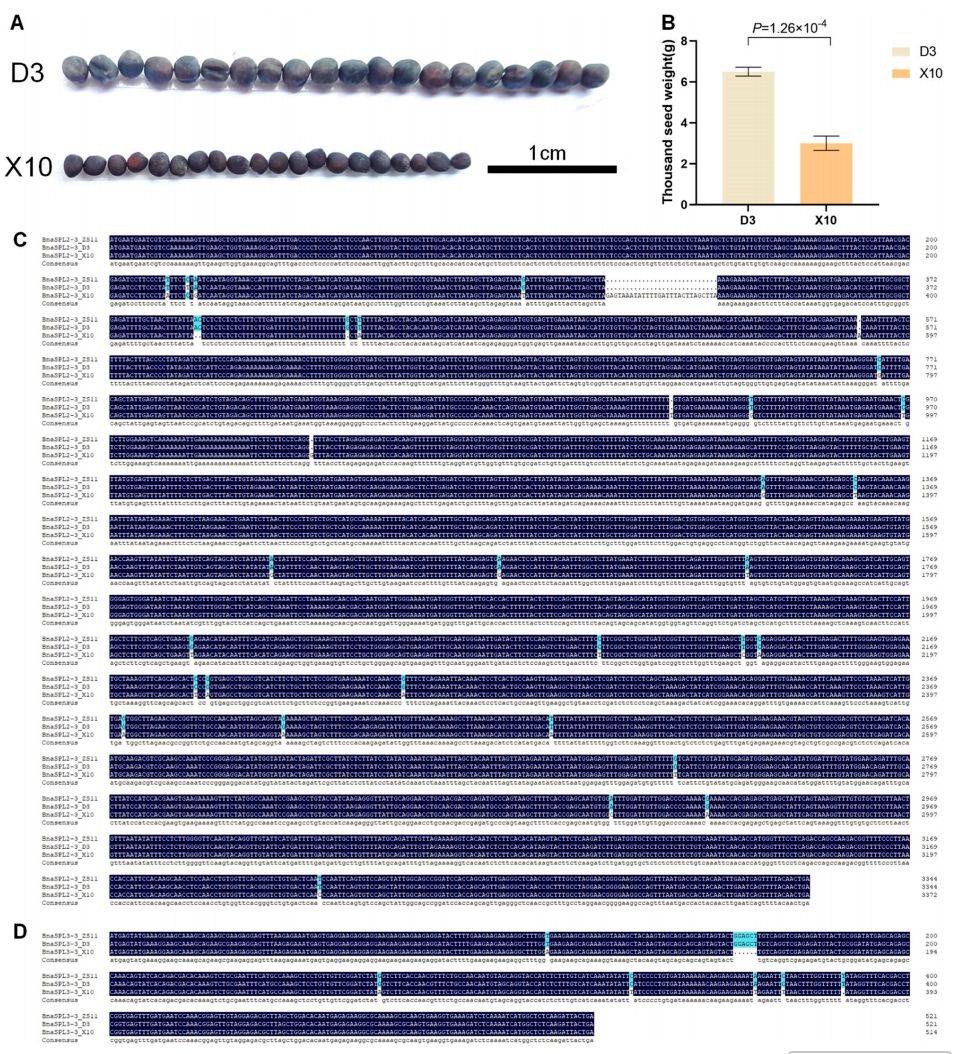
图例说明:
- A. 成熟种子大小比较: 照片直观显示大粒材料D3的种子明显大于小粒材料X10。
- B. 成熟种子千粒重比较: 柱状图定量显示D3的千粒重极显著高于X10。
- C-D. D3和X10中BnaSPL2–3和BnaSPL3–3基因序列比较: 序列比对图展示两个极端材料在这两个关键基因上的DNA🧬序列差异(SNP/InDel),验证了D3携带高产单倍型Hap1,X10携带低产单倍型Hap2

四、文章总结
本研究通过对八个甘蓝型油菜品种进行泛基因组分析,全面鉴定了SPL基因家族,共发现72个成员,并揭示了其核心、非必需和特有基因的分布。进化分析将其分为六组,并发现部分基因受到正选择。通过表达谱分析,筛选出26个在种子发育中高表达的BnaSPL基因。进一步的基因结构、顺式元件及单倍型关联分析锁定BnaSPL2–3、BnaSPL3–3、BnaSPL7–4和BnaSPL16–4为调控千粒重和产量的关键基因,其优势单倍型能稳定提升产量且抗逆性不受影响。利用极端材料的实验验证了BnaSPL2–3和BnaSPL3–3的功能。该研究为甘蓝型油菜SPL基因的功能解析奠定了重要基础,并为利用分子标记辅助选择培育高产量、高千粒重的油菜新品种提供了宝贵的候选基因和理论依据。











